 更新时间:2025-11-27
更新时间:2025-11-27 点击次数:1281
点击次数:1281

1.研究背景:
抗生素耐药病原体(尤其是多重耐药菌)的泛滥发展正威胁着全球公共卫生,无法治疗的感染所带来的潜在威胁日益显现。尽管噬菌体疗法是对抗耐药菌感染的潜在替代方案,但其疗效的有力证据仍不充分。
2.存在的问题:
噬菌体疗法面临的挑战主要源于耐药性的出现,以及其对抗细菌长期感染引发的免疫抑制状态的能力有限。细菌不仅会分泌一系列毒性因子抑制免疫细胞活性,更重要的是,它们会隐藏并改变自身抗原表位以逃避免疫细胞识别,导致感染持续进展。因此,建立噬菌体与免疫系统之间的关联,可能为现有噬菌体疗法提供突破性方向。
3.解决方法:
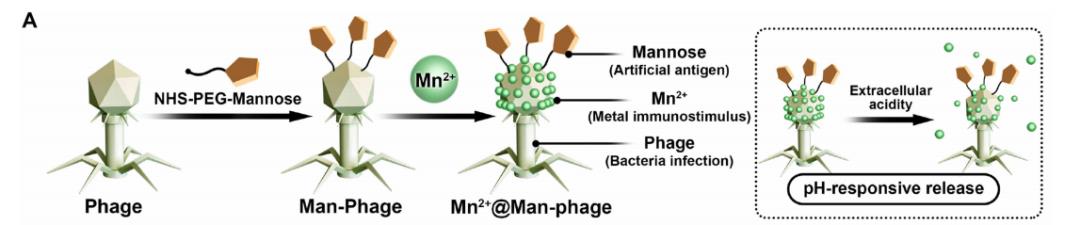
本研究开发了一种基于噬菌体的人工抗原导向免疫识别标记纳米平台(Mn2+@Man-噬菌体),该平台通过在免疫抑制微环境中搭建细菌与巨噬细胞之间的“桥梁",介导免疫级联反应的发生。
4.设计思路:
共价结合将甘露糖连接子[N-羟基琥珀酰亚胺-聚乙二醇-甘露糖(NHS-PEG-甘露糖)]修饰到噬菌体上(形成Man-噬菌体),并通过静电相互作用将锰离子(Mn²⁺)矿化到噬菌体头部。噬菌体凭借其固有生物学特性,对细菌具有*的靶向能力;而甘露糖功能化后,可通过多价结合与巨噬细胞表面的甘露糖受体作用,进一步促进免疫细胞对细菌的识别和吞噬。
实验分析-表征:
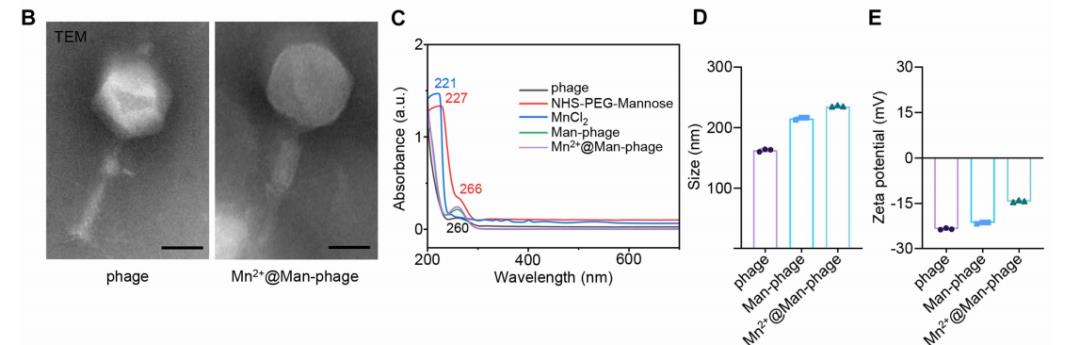
为进一步验证Mn²⁺@Man-噬菌体的表征,研究中采用TEM观察,发现制备的Mn²⁺@Man-噬菌体与裸噬菌体具有相似的规则结构,但形态发生改变(图B),且头部直径与尾长的比例与裸噬菌体存在显著差异。
紫外-可见(UV-Vis)光谱显示,Man-噬菌体和Mn²⁺@Man-噬菌体在260nm处的吸光度升高,这源于(MnCl₂)在221nm处的吸收峰以及NHS-PEG-甘露糖在227nm和266nm处的吸收峰(图C)。
动态光散射(DLS)测量显示,裸噬菌体、Man-噬菌体和Mn²⁺@Man-噬菌体的水动力学直径分别为162.70±2.29nm、215.27±2.31nm和235.2±1.22nm(图D)。相应地,经原位甘露糖化修饰和温和矿化后,Mn²⁺@Man-噬菌体的电位较裸噬菌体有所升高,但仍整体带负电,且在水溶液中具有良好的分散性和稳定性(支撑材料:图S4A、B)。
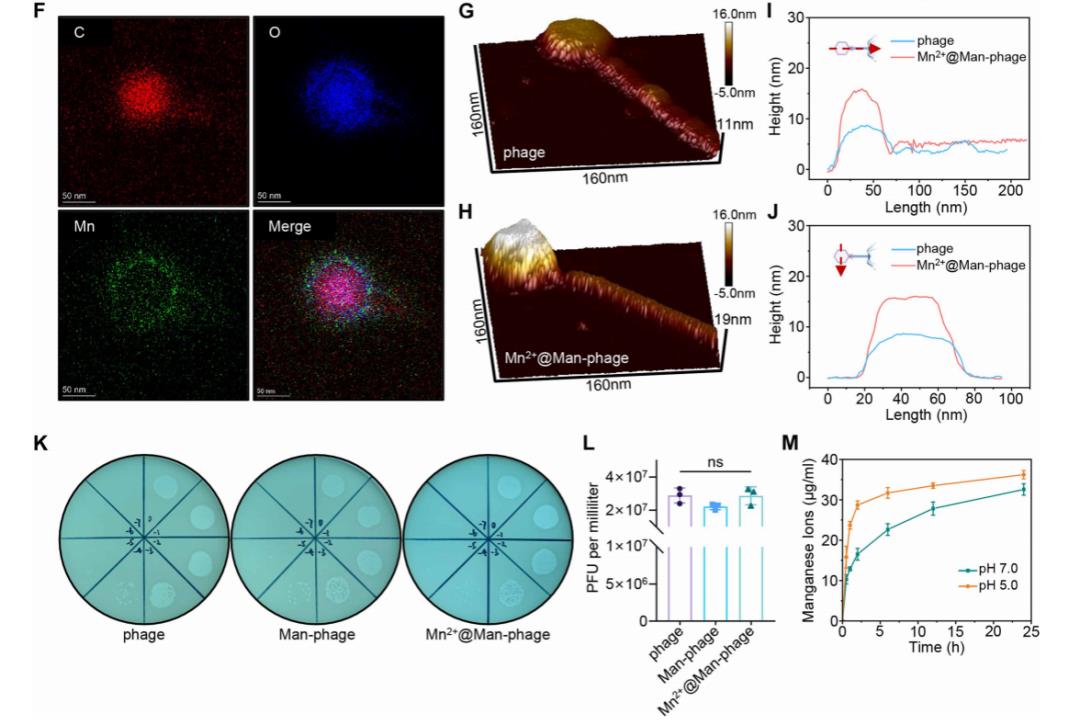
Mn²⁺@Man-噬菌体的元素 mappings显示碳(C)、氧(O)和锰(Mn)呈特征性分布(图F),与X射线能谱分析结果一致(支撑材料:图S3C),证实Mn²⁺已成功矿化到Man-噬菌体头部。通过原子力显微镜(AFM)进一步表征发现,裸噬菌体呈现典型的头-尾病毒结构(图G),而经甘露糖修饰和Mn²⁺矿化后,Mn²⁺@Man-噬菌体的整体轮廓(尤其是头部)显著增大(图H-J)。
通过噬菌斑计数评估工程化噬菌体的活性,发现Man-噬菌体和Mn²⁺@Man-噬菌体的滴度与裸噬菌体相当(图K、L),表明甘露糖化和矿化主要发生在噬菌体头部,而噬菌体尾部的完整结构得以保留。
考虑到Mn²⁺在治疗系统中关键的金属免疫调控作用,我们通过电感耦合等离子体质谱(ICP-MS)评估Mn²⁺@Man-噬菌体中Mn²⁺的释放情况:在Transwell小室外侧包裹一层10kDa透析膜以建立Mn²⁺释放模型(支撑材料:图S5A);噬菌斑实验证实,噬菌体无法透过透析膜而保留在上室,因此工程化噬菌体头部释放的Mn离子可进入下室(支撑材料:图S5B)。
如图M所示,Mn²⁺@Man-噬菌体的Mn离子释放具有pH响应性——当释放缓冲液pH从7.0降至5.0时,Mn²⁺的初始爆发释放速率显著加快。上述数据证实,我们成功合成了基于噬菌体的免疫识别标记纳米平台。
实验分析-免疫识别标记实验

将Mn²⁺@Man-噬菌体与大肠杆菌混合孵育15分钟后,再与RAW264.7巨噬细胞共培养。TEM图像显示,Mn²⁺@Man-噬菌体可锚定在大肠杆菌表面(图B),表明细菌标记成功。此外,利用异硫氰酸荧光素(FITC)标记的Mn²⁺@Man-噬菌体、1,1'-二十八烷基-3,3,3',3'-四甲基吲哚三碳菁碘化物(DiR)染色的大肠杆菌,以及表达mCherry的RAW264.7巨噬细胞,我们通过可视化观察发现,RAW264.7巨噬细胞可识别并吞噬被标记的大肠杆菌,表明Mn²⁺@Man-噬菌体能在免疫细胞与病原体之间搭建“桥梁"(图C)
流式细胞术分析显示,在Man-噬菌体介导下,RAW264.7巨噬细胞对大肠杆菌的吞噬活性高于裸噬菌体组(裸噬菌体组吞噬率为44.6%,Man-噬菌体组为56.3%),而Mn²⁺@Man-噬菌体介导的吞噬率最高(84.3%)(图D、E),荧光染色和TEM图像也证实,Mn²⁺@Man-噬菌体组的RAW264.7巨噬细胞吞噬作用较显著(图F)。

通过人工抗原评估RAW264.7巨噬细胞对被标记细菌的杀菌能力:琼脂平板培养和活/死染色结果显示,在无巨噬细胞参与时,Mn²⁺@Man-噬菌体的杀菌活性与裸噬菌体相当——与对照组相比,裸噬菌体组和Mn²⁺@Man-噬菌体组的细菌菌落数分别降至56.88±1.90%和57.45±4.67%(支撑材料:图S6A-C)。
而与RAW264.7巨噬细胞共培养24小时后,Mn²⁺@Man-噬菌体介导的细菌存活率显著降低(图G)。与对照组相比,裸噬菌体组、Man-噬菌体组和Mn²⁺@Man-噬菌体组的胞外细菌存活率分别为43.04±5.99%、29.53±1.53%和15.04±6.71%,表明Man-噬菌体可在一定程度上介导巨噬细胞清除胞外细菌,而Mn²⁺@Man-噬菌体进一步增强了这一效果。
此外,裂解细胞后收集上清液评估胞内细菌存活率,发现Mn²⁺@Man-噬菌体组与对照组的差异较显著,胞内细菌存活率仅为6.18±2.26%(图G、H)。值得注意的是,即使是噬菌体耐药菌,经纳米平台标记后,仍能被RAW264.7巨噬细胞有效吞噬和杀伤(支撑材料:图S7)。
实验分析-金属免疫调控

在各类细胞群体中,巨噬细胞的优势亚群决定了机体抗感染免疫的方向。M1型活化巨噬细胞最初被认为在对抗入侵病原体中起关键作用;而细菌长期感染会触发巨噬细胞从促炎状态向免疫抑制状态(M2型表型)转变,从而促进感染持续发展。已有研究证实,锰(Mn²⁺)等营养金属离子可激活环磷酸鸟苷-腺苷合成酶-干扰素基因刺激因子(cGAS-STING)通路,增强免疫效能。
利用Mn²⁺释放模型,分析RAW264.7巨噬细胞的极化状态:将M0型和M2型RAW264.7巨噬细胞分别在Mn²⁺释放模型的下室培养24小时。
流式细胞术分析证实,Mn²⁺@Man-噬菌体释放的Mn²⁺显著提高了M0型RAW264.7巨噬细胞中M1/M2标志物(CD86/CD206、iNOS/Arg-1)的比值。
同时,Mn²⁺@Man-噬菌体组的M2型RAW264.7巨噬细胞被有效重编程为杀菌表型:CD86⁺和iNOS⁺巨噬细胞比例分别从31.7%和3.56%升至53.2%和24.7%,而CD206⁺和Arg-1⁺巨噬细胞比例分别从82.8%和21.2%降至43.2%和10.4%(图D、E)。

上述数据表明,释放的Mn²⁺可促进M0型和M2型巨噬细胞向M1型极化。为了进一步探究Mn²⁺@Man-噬菌体释放的Mn²⁺对RAW264.7巨噬细胞中cGAS-STING通路及下游核因子κB(NF-κB)通路的影响。
Western blotting结果显示,Mn²⁺显著诱导STING、TANK结合激酶1(TBK1,促进NF-κB激活的结合蛋白)和p65的磷酸化(图F)。
研究证实,Mn²⁺可激活p65核转位,进而促进肿瘤坏死因子-α(TNF-α)、IL-6、IL-1β和IL-12等促炎细胞因子的转录和分泌(图G、H)。
这表明,在Mn²⁺@Man-噬菌体介导巨噬细胞识别和吞噬细菌的过程中,释放的Mn²⁺可通过激活cGAS-STING-NF-κB通路,将巨噬细胞重编程为M1型,进一步增强其抗菌能力。
实验分析-转录组分析
深入探究Mn²⁺@Man-噬菌体诱导的基因组变化,采用RNA测序分析其转录组图谱。
火山图显示,Mn²⁺@Man-噬菌体处理后共鉴定出2200多个差异表达基因(DEG),且这些基因主要参与促炎应答(图A)。KEGG富集分析显示,Mn²⁺@Man-噬菌体处理后,免疫相关激活通路中候选基因的富集因子变化较为显著,包括IL-17、TNF、NF-κB和核苷酸结合寡聚化结构域(NOD)样信号通路(图5B)。基因集富集分析(GSEA)结果与之一致,进一步证实RAW264.7巨噬细胞对病原体的激活效应(图C)。
转录图谱热图显示,Mn²⁺@Man-噬菌体处理后,巨噬细胞中免疫激活相关基因表达上调,包括抗原呈递共刺激分子(如CD40、CD80、CD86、Icosl)和促炎细胞因子(如IL-6、IL-1β、Nos2、TNF-α、IL-12)的转录水平广泛升高(图D)。此外,实时定量聚合酶链反应(qPCR)检测也发现,促炎基因(Tnfa、Il1b、Nos2)的表达呈显著上调趋势(图E)。
此外,基因本体论(GO)数据库分析显示,上调基因与巨噬细胞表型和金属离子结合相关——生物学过程和分子功能的顶级富集术语包括巨噬细胞激活、促炎应答、Mn²⁺结合以及免疫应答相关细胞因子产生(图F)。

实验分析-靶向潜力探究


为探究Mn²⁺@Man-噬菌体的感染靶向潜力,在单只小鼠上进行了对比建模实验:右侧胫骨植入无菌物体,左侧胫骨植入被大肠杆菌污染的植入物(图A)。大肠杆菌感染24小时后,向小鼠静脉注射裸噬菌体和Mn²⁺@Man-噬菌体,并利用活体荧光成像系统(IVIS)检测其分布。
如图B、C所示,注射后2小时即可在感染胫骨处观察到荧光信号,且信号强度随时间逐渐增强,在注射后12小时达到峰值;而未感染部位的荧光信号无显著变化。这些结果表明,Mn²⁺@Man-噬菌体可精准结合大肠杆菌,从而实现细菌清除和免疫调控。
此外,我们评估了Mn²⁺@Man-噬菌体的治疗效果(图D)。治疗后第7天,接受Mn²⁺@Man-噬菌体治疗的小鼠感染严重程度显著减轻:苏木精-伊红(H&E)染色和吉姆萨染色显示,骨髓中中性粒细胞浸润和细菌载量显著降低(图E);扫描电子显微镜(SEM)结果显示,Mn²⁺@Man-噬菌体组植入物上的细菌菌落被显著清除(图E)。
同时,膝关节肿胀减轻、膝关节周长缩小、体重恢复以及组织学评分降低,均表明Mn²⁺@Man-噬菌体具有优异的抗菌效果(图F-I)。
琼脂平板培养结果与上述一致:Mn²⁺@Man-噬菌体处理后,骨髓和植入物中的细菌菌落数均大幅减少。与对照组相比,Mn²⁺@Man-噬菌体组骨髓和植入物中的细菌计数均降低6个数量级,而裸噬菌体组仅降低4个数量级(图J)。
实验分析-免疫机制

免疫荧光染色图像显示,Mn²⁺@Man-噬菌体可介导巨噬细胞向M1型极化,表现为CD86和iNOS表达升高(图B、C)。
M1型极化巨噬细胞在宿主抗感染防御中起重要作用:它们迁移至感染部位后,识别并吞噬入侵细菌,随后发挥细胞毒性作用,并向适应性免疫细胞呈递抗原。同时,CD4⁺和CD8⁺T细胞的级联激活需要M1型巨噬细胞提供充足的抗原呈递。
Mn²⁺@Man-噬菌体治疗后,免疫组化染色显示骨髓中CD4⁺和CD8⁺T细胞浸润增加,表明Mn²⁺@Man-噬菌体激活的巨噬细胞可通过吞噬细菌并向T细胞呈递抗原,促进T细胞激活(图D、E)。

此外,通过流式细胞术全面分析了骨髓中的免疫细胞组成及免疫图谱变化:与上述结果一致,Mn²⁺@Man-噬菌体治疗显著诱导骨髓中的巨噬细胞向杀菌表型转变(图F、G);Mn²⁺@Man-噬菌体组中iNOS⁺F4/80⁺细胞比例升至10.7%(对照组为4.62%),而Arg-1⁺F4/80⁺巨噬细胞比例降低约一半(图J)。
Mn²⁺@Man-噬菌体治疗后,感染部位CD4⁺和CD8⁺T细胞比例显著增加,表明适应性免疫应答被激活(图H、I);且Mn²⁺@Man-噬菌体显著增强T细胞活性——IFN-γ⁺CD4⁺和IFN-γ⁺CD8⁺T细胞比例分别是其他组的4倍和3倍(图K)。胫骨骨髓悬液的酶联免疫吸附试验(ELISA)结果显示,Mn²⁺@Man-噬菌体组中TNF-α、IL-12和IFN-γ水平显著升高,而IL-10水平相应降低(图L)。
5.研究结论
本研究基于噬菌体捕食的固有机制,提出了一种创新的抗细菌感染噬菌体免疫疗法——该疗法通过在宿主免疫抑制微环境中重建细菌与巨噬细胞之间的“桥梁",介导抗感染免疫级联反应的发生。所有数据均证实,我们的纳米平台可有效激活免疫应答,实现细菌清除。此外,研究结果表明,经修饰的噬菌体可作为细菌表面的“人工抗原",用于宿主细胞免疫监视;且该治疗系统可扩展应用于多种免疫细胞和病原体。
鉴于传统抗生素替代方案的迫切需求,本策略通过整合免疫治疗特性,为噬菌体疗法带来创新,为其在临床感染治疗中的应用奠定基础。
6.创新点
机制创新:揭示噬菌体单独疗法的局限:不能有效激活宿主免疫,导致耐药菌和免疫抑制环境持续存在。提出通过“人工抗原标记"重建细菌与免疫系统的连接。
材料策略创新:将噬菌体残留衣壳“二次利用"为人工免疫标记载体。引入 甘露糖+Mn²⁺ 双功能化:甘露糖增强巨噬细胞受体识别。Mn²⁺作为免疫金属因子,触发 cGAS-STING 信号,推动 M1 极化。
治疗模式创新:不再仅依赖噬菌体直接裂解,而是通过“噬菌体-宿主免疫双桥梁"实现细菌清除。同时激活 先天免疫(巨噬细胞)+ 适应性免疫(T细胞),构建系统性免疫反应。
该策略可适配多种噬菌体,不限于单一病原体,甚至可与噬菌体鸡尾酒联合。可接驳不同免疫配体,具备较高的可扩展性与转化潜力。

 当前位置:
当前位置:
